

- Jointly published in the American Bee Journal and in Bee Culture, October 2012.
Also at eXtension.org: http://www.extension.org/pages/65450/varroa-mite-reproductive-biology
Zachary Huang, Michigan State University
The varroa mite (Varroa destructor Anderson and Trueman) is an ecto-parasite of the Western honey bee (Apis mellifera) and is distributed worldwide. Because A. mellifera colonies almost always die within two to three years after mite infestation, if not treated, feral bee colonies (unmanaged colonies in the wild) in U.S. were almost totally wiped out by this mite around 1995, less than a decade after it was introduced to the USA (around 1987). There is anecdotal evidence that honey bees might be becoming feral again in recent years (resistant genetics possibly leaking out due to swarming), but there is no systematic study proving this. Unless otherwise noted, throughout this paper I will use “varroa”, “varroa mite” or the generic “mite” interchangeably to refer to V. destructor. The varroa mite is currently the most severe pest of managed honey bees worldwide. Understanding the varroa mite’s reproductive biology will therefore allow us to better manage this important pest.
The Life Cycle of Varroa
Varroa mite life cycle has two stages (Fig. 1). During the phoretic stage, mites ride on adult workers or drones, at the same time feeding on blood (hemolymph) from bees, usually from the inter-segmental membrane on the abdomen. The phoretic stage lasts about 5-11 days when there is brood in the colony. Of course, mites are forced to remain phoretic if there is no brood, and this can last 5-6 months in cold climates. Mites change hosts (hop from one bee to another) often and this contributes to transmission of various viruses, by picking them up from one bee and inject to another during feeding. Mites experience higher mortality during the phoretic stage, because they make mistakes, and fall to the screen bottom board, if a hive has one, get bitten by workers during grooming, or die due to old age. The “natural drop” on a screened bottom board reflects a combination of all of these factors. However, the total of these fallen mites are less than 20% of the population. Therefore, using a bottom-screen-board alone will reduce, but not eliminate, chemical use for varroa management. The phoretic stage is important for mites to transfer horizontally to other colonies, by being accidentally dropped onto flowers and then picked up by other foragers (this probably does happen, but we do not know the actual probability), by mite-carrying bees drifting to another colony, or finally by bees robbing a colony dying from mite infestation. In the last situation, we are actually selecting for mites with high virulence, because while in a natural forest, mites that kill a colony will also die with their host (due to the low likelihood of being found by a neighboring colony), while in an apiary this robbing behavior is guaranteed, insuring the successful transfer of mites from the dying colony to another, where it will repeat the cycle again.
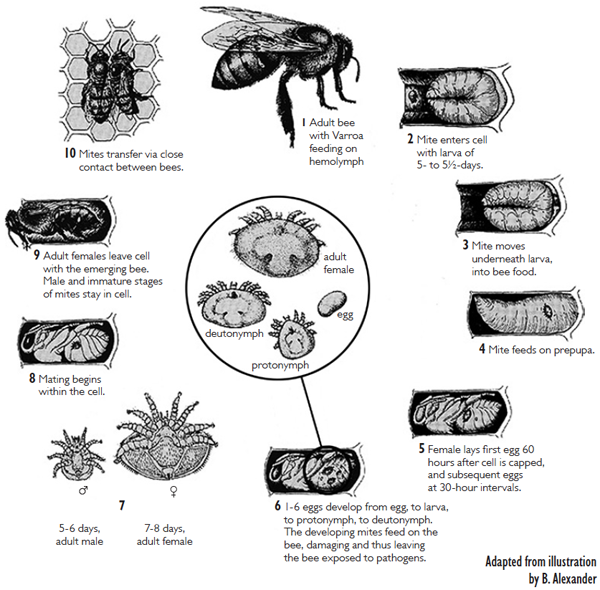
Fig. 1. Varroa destructor life cycle, adapted from B. Alexander
The other stage is the reproductive stage, and only during this time when it is possible for mites to increase their population. This occurs only under the capped brood cell. Mature female mites are already mated when they emerge, as bees emerge from the cell. The varroa mite invades a host (worker or drone larvae) cell just prior the cell being capped. Once inside, she will hide in the brood food in an upside-down position (viewed from the top of the cell). Mites have special appendages called “peretrimes” (essentially as snorkeling tubes) that help them breathe. Shortly after a cell is capped, the larva inside will spin a cocoon, and then becoming a prepupa. The mite will not feed until about five hours after the cell is capped (after spinning is done) and the first egg is laid 70 hours after cell capping. The first egg is not fertilized, and becomes a male. This is the same as in honey bees, as both organisms have what is called the “haplodiploidy” sex-determination mechanism, whereby males are haploid (having no father) and females are diploid (having both parents). After this, approximately every thirty hours, the mite lays a female egg. If the mother mite was not mated properly, then all of her offspring will be males. A total of five (on worker pupae) or six eggs (on drone pupae) can be laid in a capped cell. However, because worker bees will emerge about eleven days after capping, and drones fourteen days, but a daughter mite takes six days to mature , most of these eggs do not have time to develop into adults, (6 + 70 hrs delay in egg laying + 1 day for first egg as male=10 days, leaving only 1 daughter to mature). The males and the unsclerotized (white) females, who are not fully developed, all die shortly due to dehydration after a cell is opened (after bee emergence, or uncapping by hygienic bees). Therefore only the mature, tanned female mites,, but not most nymph stages and males, are seen by most beekeepers (Fig. 2). Males will mate with a female repeatedly to result in a total of about 35 spermatozoa inside the female spermatheca.
Varroa mites have “fecal sites” on the cell, where they deposit their feces, which are white due to a high concentration (~95%) of guanine. For some unknown reason, any mite that defecates on the pupa directly are also sterile (Fig. 3).

Fig. 2. Mature, immature females and mature males of Varroa. Clockwide from top left: mature daughter mite, mother mite, two mature males and an immature (deutonymph) daughter. A younger stage (protonymph) of female is not in this photo.

Fig. 3. Sterile mites with defecation on bee abdomen. All these six mites did not reproduce and all had defecated on the Apis cerana workers. This is also true in Apis mellifera: if a mite has defecated on the pupae, she would have no daughters. If she has daughter mites, she would be defecating on the wall.
Methods for Studying Mite Reproduction
There are two methods for studying mite reproduction. One method is simply to survey, uncapping worker or drone cells in colonies and determining the percentage of mites that reproduced (fertility), or the number of offspring (fecundity) of mites. This method gives information about what is happening under natural conditions, but the information one gets is limited because nothing is controlled or manipulated. Another method is to perform manipulations, either on the mites or on the host, then artificially introduce mites into the cells and wait 9-10 days to determine fertility and fecundity. The frames can be reintroduced into a colony (which risks removal by bees due to hygienic behavior), or incubated in a laboratory. The basic steps for this process are as follows. 1). Harvest mites either from brood cells or from phoretic hosts in a colony with a high number of mites. We now routinely harvest mites using the sugar dusting method,, and then clean the sugar off them using a moistened brush. 2). Obtain brood cells that are recently capped (within 6 hours). This should be from a colony with no mites, so that you do not have natural invasion into the cells that you are trying to introduce mites to– if you have two mites in one cell, you do not know which one is the introduced one. Mites will not reproduce if introduced into cells that have been capped for more than 14 hours. Ideally, the cells should be capped within the last six hours. Scientists often use a piece of transparency to map the brood; marking those that are being capped (with holes on the cap), waiting for six hours, coming back and marking again. Those cells that had a hole the first time, but were totally capped after, are the cells that you need. Make sure to mark the two holes (from fixing the transparency on the frame) on the wooden frame with Sharpies also, because bees may seal the small holes with wax, and you lose the reference. 3). Open the cell slightly using a fine scalpel, an insect pin, or a pair of fine forceps, and introduce a mite carefully into the small slit using a horse hair or a fine brush. 4). Push the wax capping back, and seal it with melted beeswax with a brush. 5). Keep the frame upright at all times, and the relative humidity at 50%, and at a temperature of 32-35 °C. 6). Check the frame daily for signs of wax moth larvae, because they can destroy the data; do not put the frame flat on a surface, but keep it upright during this process. 7). Finally, on day nine (mite transfer day designated as day one), one can uncap the cells and count and record how many males, daughter mites (mature, deutonymphs and protonymphs) are there.
Host Preference During Phoretic Stage
Whether or not varroa mites can choose phoretic hosts was studied previously, using caged bees and petri dishes. Varroa mites preferred nurses when they were presented with a choice between foragers and nurses in a petri dish. Mites also transferred more often to young bees than to old bees, when they were confronted with freshly frozen young and old bees. This discrimination by varroa was later shown to be related to the repellent effect of geraniol, a component of the Nasonov pheromone, which is high in foragers. However, it was not clear whether mites show the same preference under a more realistic colony condition. One study showed nurses had a higher percentage of mites than newly emerged bees, but no difference was found between nurses and foragers. Another study found nurses were the most preferred but the experiment was conducted in one colony (i.e. not replicated). My laboratory studied mite distribution among one-day-olds, nurses (5-11 day old marked bees recovered from a colony) and foragers (unknown age but the average age of foraging bees should be higher than 21 days in a typical colony), and found a clear preference of nurses > day-old-bees > foragers (X. Xie, Z.Y. Huang and Z. Zeng, in preparation). Thus, mites do show the same preference for nurses, even in a colony setting.
Why Phoretic Stage?
Scientists were puzzled as to why mites bother to go through a phoretic stage, because they experience a high mortality rate during this period. Under laboratory conditions, varroa can reproduce successfully without a phoretic stage. That is, mites that were transferred immediately upon bee emergence to another newly-capped brood cell still reproduced, for up to seven cycles. The average number of total offspring (including males) was four during the first four to five cycles. This seemed higher than in our experiment (Fig. 4). However, upon closer inspection, the author said there were mites that did not have offspring (21.7%), and that this was most likely not included in the calculation. So the actual fecundity was 4*(1-0.217) = 3.17, which also included the males. After subtracting the males (which were about two per mother, instead of one, strangely, in their study, Fig. 4 of De Ruijter, 1987), we have 1.17 female offspring per mother. This would be slightly lower than the 1.6 female offspring per mother that we observed in mites fed on newly emerged bees. Therefore, mites that do not experience the phoretic stage have a lower fertility, especially compared to those hosted by younger nurses (see below).
Our recent study showed that mites preferred nurses, perhaps not only because of their proximity to larvae (nurses inspect/feed larvae frequently), but also because nurses provide phoretic mites extra nutrition for reproduction. Our study found that mites artificially fed on nurses had the highest number of offspring, followed by those on foragers, and those fed on the newly-emerged bees having the lowest number of offspring. In addition, when we compared the fecundity of mites hosted by bees with different ages, we found a significant negative relationship between mite fertility and the age of nurses (Fig. 4).
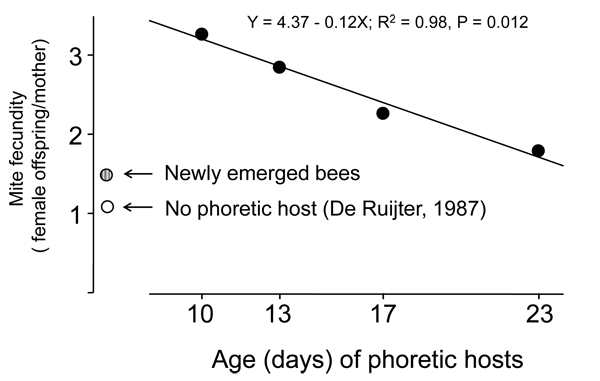
Fig. 4. Negative relationship between mite fecundity and age of phoretic hosts. This relationship is only true if one uses bees of nurse ages and older, because newly emerged bees as phoretic hosts also cause low fecundity (show by the grey circle), which was slightly higher than those without going through a phoretic host (empty circle, data calculated from De Ruijter, 1987).
Differences in Mite Reproduction is Important for Resistance Against Mites
Varroa mites can reproduce on both worker and drone brood in Apis mellifera, but reproduce exclusively on drone brood in A. cerana, its original host. Many factors, such as grooming behavior (removing mites during phoretic stage from adults), hygienic behavior (removing mites from pupae during reproductive stage), duration of brood state, and attractiveness of brood, contribute to varroa tolerance (reviewed by Büchler, 1994). However, I think that reduced reproduction (including both reduced fertility and fecundity) on worker brood is the most significant factor for honey bee resistance against the Varroa mite. This is because the infertility of Varroa on worker brood correlates well with the degree of tolerance of that bee to the mite. For example, A. cerana is highly tolerant to the mite and causes 100% mite infertility in worker brood; the Africanized bee (A. mellifera scutellata) is intermediately tolerant with a 40% infertility; while A. mellifera in U.S. is the least tolerant with the lowest infertility rate (10-20%) in worker brood. In a strain of European bee that was artificially selected to be tolerant of mites, infertility of mites plays the most significant role in depressing the mite population, while other factors (such as grooming behavior, hygienic behavior, and the duration of the postcapping period) are not as important (Harbo and Hoopingarner, 1997). Although we currently know that the original “SMR” (suppressing mite reproduction) trait is actually due to “VSH” (varroa sensitive hygiene), VSH can be considered a special trait causing lower reproduction, due to the interruption of the reproductive cycle of the mites, especially since the bees do not open cells containing non-reproducing mites, but rather target those having mite daughters.
Factors Affecting Mite Reproduction
A. Effect of Caste of Brood
It has been known for a long time that varroa mites preferred drone brood over worker brood, in a ratio of nine to one. That is, if there is an equal number of cells available, the drone brood would harbor nine times as many mites as the worker brood. Natural selection undoubtedly favored mites that preferred drones, because drone brood has a longer capped-period, enabling more daughter mites to mature. Indeed, Martin (1994, 1995) calculated the effective reproduction rate (i.e. the number of vial/mature daughters per invading mother) as 1.3–1.45 in a single infested worker brood, while for drone brood it was 2.2–2.6. In A. mellifera, transferring mites from drone to worker brood always deceased mites’ reproduction rate, while transferring mites from worker to drone brood increased reproduction rate. Queen larvae would be a dead end for invading mites, because queens emerge at 16 days, five days faster than a worker, thus leaving the daughter mites no time to mature. Varroa mites do avoid queen cells, apparently due to some chemical odor from royal jelly.
B. Effect of Host Species
The transferring of mites across different species suggests that host species also affects mite reproduction. When mites from A. cerana were introduced to A. mellifera worker brood, only 10% of the mites reproduced, while 80% reproduced when A. mellifera mites were transferred to A. cerana worker brood. In our study, Varroa destructor, Korea haplotype, from A. mellifera reproduced equally well (all > 90% reproduced), regardless of whether it transferred to A. mellifera or to A. cerana, in both drone and worker castes (Ting Zhou, Shuangxiu Huang and Zachary Huang, unpublished data). In contrast, V. destructor, Vietnam haplotype, from A. cerana only reproduced on A. cerana drones (83% reproduced, N=62), and not on A. cerana workers (0% reproduced on workers, N=60). These results suggest that the mites on the two honey bee species are different: mites from A. cerana refrain from reproducing on worker brood of the same species, and mites from A. mellifera reproduce well on worker brood, regardless of the host species. It appears that only the Korea haplotype of V. destructor had a genetic change that enabled it to reproduce on either drone or worker brood in A. mellifera, therefore allowing it to build up to levels damaging to the bees. In China, my colleagues and I did not find damaging levels of V. destructor in A. cerana colonies – in fact, in most locations, the mites could not be found. When we found it, it was the Vietnam haplotype which does not reproduce in the worker brood of A. cerana. It is not clear why the Korea haplotype of V. destructor does not cause damage in A. cerana, since they can reproduce in both worker and drone brood in transfer experiments. However, it is possible that they do not reproduce on worker brood under natural conditions, when both the phoretic and reproductive hosts were A. cerana. Thus, transfer experiments should be supplemented with observation under natural conditions for the full picture.
C. Effect of Cell Size
Partly because mites reproduce better in drone brood than worker brood, people tend to think that smaller cells would decrease mite reproduction. However two recent studies show that there was either no difference in mite population between colonies (Ellis et al., 2009) using “small cells” (4.8 to 4.9 mm diameter) and regular foundations (5.2-5.4 mm), or small cells actually had a significantly higher mite population (Berry et al., 2010). Unfortunately, neither of these recent studies determined the fecundity or fertility of mites in the two types of cells.
Earlier studies were conflicting. Taylor et al. (2007) found that “foundation” cell size did not affect the reproductive success of V. destructor, but more mites invaded cells drawn from the 4.8mm foundation . However, Piccirillo and De Jong (2003) and Maggi et al. (2010) found that mite invasion rate increased positively, and linearly, with the width of worker and drone brood cells, probably because brood that develops in large cells receive more visits from nurses, increasing the invasion chance. Maggi et al. (2010) also found that the percentage of fertile mites was lower in smaller cells. An earlier study (Message and Goncalves, 1995) showed in Africanized bees, larger cells had a higher invasion rate, and also had higher effective fecundity in mites.
Our own study suggests that cells that are too large also reduce mite reproduction (Zhou et al., 2001). In a study trying to determine the mechanisms of why varroa mites do not reproduce on worker brood of A. cerana, we accidentally discovered that in both A. cerana and A. mellifera queens laid worker eggs in drone cells in the fall. We took advantage of this, and compared the reproductive output of mites on two hosts: workers reared in worker-cells (WW) or workers reared in drone-cells (WD). In 2001, both the fertility and fecundity of the two groups were significantly different (Fig. 4). It is not clear why mites would reproduce less on identical hosts that were housed in larger cells. One possibility is that workers reared in drone cells are fed a different diet by nurses (One study showed workers reared in drone cells were heavier and had more ovaries, suggesting a different diet or more nutrition). A second possibility is that workers spin larger cocoons in drone cells, and mites detect the extra space, and this affects their reproduction.
D. Effect of Humidity
Kraus and Velthuis (1997) wondered why varroa mites were not as big a problem in the tropics (besides that fact that most bees were African), and tested in the laboratory to see if high relative humidity would inhibit mite reproduction. They artificially transferred single mites into newly capped cells, and then kept the brood in an incubator. When relative humidity (RH) was set at 59–68%, on average, 53% of the mites produced offspring (N=174 mites); under 79–85% RH, only 2% (N = 127) of the mites reproduced. The difference in mite fertility was highly significant. My postdoctor recently incorrectly set the incubator at a RH of 75% (instead of 50%), and very few mites reproduced as a result. If there are ways to artificially increase the hive RH to about 80%, then the varroa mite population will never increase to a damaging level.
E. Effect of Comb Movement
Aside from where they defecate, varroa mites are also very picky about where they feed. The mother herds her “children” to one particular feeding site on the pupa (between the pair of hind legs on the ventral side of the abdomen), and then leads them back to the defecation site. Therefore, any rotation of combs will cause the movement of the host pupa? and perhaps causes disorientation of the mites. The “Kônya beehive with rotating frame [sic] of brood nest” was invented (and patented) by Lajos Kônya, from Hungary. The hive body has round frames and they rotate ten degrees per hour, thus completing a circle in 36 hours. This is powered by a 12 volt battery. Varroa mites are not able to reproduce, due to the constant rotation of the cells. I was pretty confident that the claims were true based on mite reproductive biology. However, an abstract (Aumeier et al., 2006) said they studied the rotation of combs on mite reproduction for three years and found no evidence that it worked. Daily rotating or shaking of brood cells neither “affected fertility (93-100%) nor fecundity (2.6-3.0) of reproductive mites or mortality of mite offspring in the brood cells.” This is a bit surprising because I thought prior to filing for the patent, the inventor should have obtained data showing that the rotation affected mite reproduction? However the study did report that swarm cells were removed due to the rotation, so the Kônya hive does work for swarm prevention.
F. Effect of Host Age, a Kairomone, a Hormone, a Pheromone, and Genes
Varroa mites that have been artificially introduced into brood cells that have been capped for over 14 hours will never reproduce. Of mites that were introduced to cells 12 hours post-capping, about 10% reproduced. Garrido and Rosenkranz (2004) therefore hypothesized that an odor from fifth instar larvae are used as signals by mites to activate their ovaries. This chemical, since it benefits the receiver, should be called a kairomone. They then designed a special cage to confine mites over various testing objects, and found that mites activated oogenesis after perceiving larval volatilities, and those mites were deprived of food, since any bee blood could also contain signals. Pentane extracts of the larval cuticle also caused ovary activation, suggesting that the chemical signal is polar. The chemical remains unidentified.
Initially there was a hypothesis that the juvenile hormone (JH) in the honey bee larvae/pupae could be the factor that activated varroa ovaries, and therefore regulated their reproduction. JH is an important hormone and in most insects it regulates oogenesis and spermatogenesis. This theory was abandoned after observing no differences in JH titers in Africanized and European bee larvae, even though it has been proven that Africanized bees have much lower mite reproduction rates (mainly due to a much higher percentage of infertile mites).
When more than one mite invade a single brood cell, the per capita fecundity decreases, as the number of mother mites per cell increases. Mites invading brood cells in older combs also have fewer offspring. This led scientists to speculate that mites themselves might have a chemical to inhibit each other’s reproduction (a pheromone). A chemical, (Z)-8-heptadecene, was identified. In the laboratory, it caused a 30% reduction in mite fecundity. When tested in the colony, the average number of offspring was 3.48 in cells treated with (Z)-8-heptadecene, but 3.96 in control cells. This difference was small, but statistically, highly significant (P < 0.01). The effective fecundity (number of potentially mated daughters) was 0.94 in treated cells, and 1.31 in control cells; and this level of difference should have a rather large impact on population growth.
To initiate reproduction, many complicated physiological processes have to be in place. Finding genes critical to these processes can potentially lead to new ways of mite control. My lab recently started a project to hunt for genes important for survival and reproduction in mites, through the use of RNA interference (RNAi). RNAi is a method to inject a relatively large stretch of double stranded RNA (400-500 base pair long), which gets cut into 20-30 bases long, then binds to some complexes which eventually finds complementary stretches of RNA and degrade them, resulting in the reduction of a targeted gene’s messenger RNA, ultimately their protein product. Our basic principle is to search for the same genes regulating survival or reproduction in related organisms (e.g. ticks) in the mite genome, synthesize double stranded (ds) RNA, inject the dsRNA into mites, and then observe their survival. If the injected mites survive, then we proceed to observe their reproduction by introducing them into newly capped brood cells. Once a list of genes are found, we then need to ensure that the dsRNA are specific to mites, and will not affect bees, then find a way to introduce the dsRNA to mites (either directly or to the hemolymph of bees, which then get passed to mites due to their feeding).
Summary
In summary, many factors can affect mite reproduction. These range from type of reproductive host (drone, worker, or queen), cell size, age of the larvae, phoretic host type, relative humidity, or even movement of the combs. The more we understand about how reproduction is regulated in mites, the easier it will be for us to find a way that disrupts mite reproduction while not harming the bees. The trick is that the method has to be easy and economical to implement. Thus, “basic research” into the reproductive biology of mites will eventually become useful to beekeepers, as it may one day provide a new method for mite control.
Acknowledgement
I thank Melissa Huang, Meghan Milbrath, and Xianbing Xie for reviewing this manuscript. Research in the author’s laboratory on varroa reproduction was supported by a USDA SCP grant, the Chinese Natural Science Foundation, MSU GREEEN, and more recently (RNAi project) by grants from the Foundation for the Preservation of Honey Bees, National Honey Board, Almond Board of California, and MSU GREEEN. Research on Nosema was supported by a Managed Pollinator CAP USDA NIFA 20098511805718.
Main References
- Anonymous, KÔNYA’ beehive with rotating frame of broodnest http://www.imkerei.com/articles/us/rotating_broodnest.htm. visited July 20, 2012.
- Büchler, R., 1994. Varroa tolerance in honey bees – occurrence, characters and breeding. Bee World 49: 6–18.
- De Ruijter, A., 1987. Reproduction of Varroa jacobsoni during successive brood cycles. Apidologie 18: 321–326.
- Rosenkranz, P., P. Aumeier, B.Ziegelmann. 2010. Biology and Control of Varroa destructor. J. Invert. Pathol. 108: S96-S119.
- Zhou, T., J. Yao, S.X. Huang, Z.Y. Huang. 2001. Larger cell size reduces varroa mite reproduction. Proceedings of the American Bee Research Conference, American Bee Journal 141: 895-896.
This article was written in 2012. Since then significant findings have been made, the most notably that mites do not feed on hemolymph only, but perhaps fat+hemolymph for both their survival and reproduction. I also fixed the images since the original images did not show correctly. Zach, updated Feb 21, 2021.
Varroa have been proven to feed on fat bodies not hemolymph. Time to update . Thanks for your help.
Yes, this version was written before the Ramsey paper.