Dose Response of Honey Bee Behavioral Development to Nosema Apis
Ron Huarong Lin and Zachary Huang
Department of Entomology, Michigan State University, East Lansing, MI 48824
Introduction
Nosema apis is a microsporidian parasite of honey bee. It causes nosema disease, which is a major threat to the beekeeping industry. To devise more effective way to control Nosema apis. We need to learn more about the mode of action of Nosema in worker honey bees. Nosema infection induces earlier worker orientation and foraging, and earlier degeneration of hypopharyngeal glands. These symptoms are similar to the effects of juvenile hormone on worker behaviour and physiology. Nosema infection causes extra larval molts in other insects, Tribolium castaneum, possibly due to the JH or JH-like chemical produced by Nosema whitei (Fisher and Sandborn, 1964). These facts suggest that the effects of Nosema on honey bee workers might be linked to JH production by Nosema or related to Nosema infection.
Previous work suggested that bees from different colonies responded to Nosema infection differently (Huang, unpublished). We undertook this study to investigate the mode of action of Nosema on bees, first by screening bees sensitive to Nosema infection and then testing dose response of the worker behavioral development to nosema infection.
Experiment 1. Screening bees sensitive to Nosema infection
Materials and Methods
Bee source colonies: package bees were purchased from Georgia, headed by queens from Kentucky, Georgia and California in early spring, 1999. In June-July, 1999, sealed brood from these grown colonies were removed and separately incubated in holding cages overnight. Next morning newly emerged bees were marked on their thoraces with enamel paint (Testor's PLA).
Nosema inoculation: newly emerged bees were individually fed with 1 ml 50% sugar syrup containing no spores as control or containing 4,500 spores as treatment, with each group consisting of 50 or 100 bees. Individual workers were isolated for 30 min to eliminate spore transfer. Then bees were introduced into a natural colony, in which 2 Apistan strips were applied to control varroa mites to avoid any differences which may be caused by the the mite parasitization.
Sampling bees: after bee introduction, marked bees returning from flight were collected in front of the entrance daily, 1 hr in the morning and 1hr in the afternoon until 50% of marked bees in a group were recovered. We referred bees returning from orientation and foraging flights as foragers in this study.
Statistic analysis: G-test was performed to analyze forager data.
Results
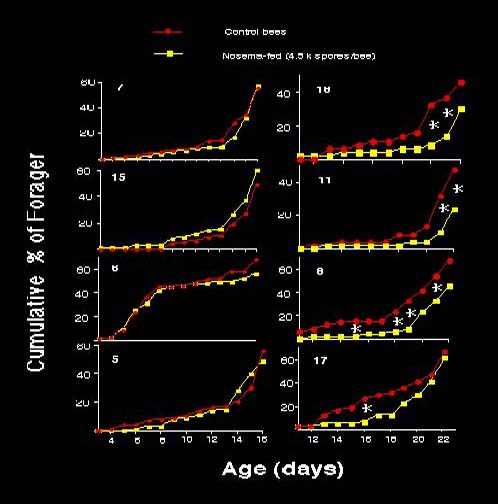
Nosema infection increased the proportion of bees to become foragers in 4 colonies (8, 11, 16 &17, see Fig. 1), but did not affect the other colonies (5, 6, 7 and 15.).
There was no cross contamination of spores among control and infected workers in the same colonies because no spores were found in control bees at the end of experiment.
Fig. 1. Effect of Nosema infection on the likelihood of becoming foragers. In June, bees from Colonies 8, 11, 16 &17 infected with Nosema showed a significantly higher proportion of foragers than control bees when bees were 15-23 day old. In July, infected workers from colonies 5, 6, 7 and 15 had the similar proportion of foragers as control bees. An asterisk (*) between the control and treatment indicates a significantly different proportion of foragers between the two groups on that day.
Experiment 2. Dose response of worker behavior to Nosema infection
Materials and Methods
After obtaining bees sensitive to Nosema, colonies 11 and 17 were chosen as the source colonies. Newly emerged workers from each colony were divided into 4 groups (100 bees/group). Bees from each group were individually fed with 1 ml of 50% sugar syrup containing 0, 440, 4,400 or 44,000 spores. Bees were painted a different color for each group. After 30 min isolation, all bees were introduced into the same colonies. Marked bees were collected daily starting the 3rd day after bee introduction.
When bees were 10, 15 and 20 days old, 10 marked bees from each group were collected inside a hive, and kept cold-anesthetized and bled. Samples of (2-10ml) hemolymph in 500ml acetonitrile were stored in -80 degrees C until analysis for JH titer.
Results
Workers from two sensitive colonies responded to Nosema infection differently. In colony 11, only the highest dosage at 44,000/bee induced more foragers than the control. In colony 17, bees responded in a dose-dependent manner to different spore concentrations (Fig. 2).
Analysis of JH is still underway.
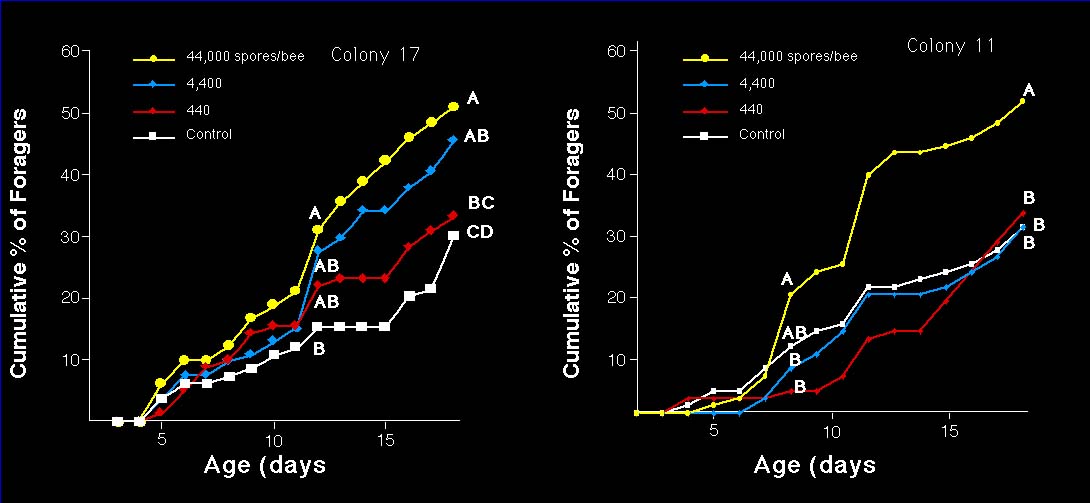
Fig. 2. Dose response of worker behavioral development in response to Nosema. In colony 11, bees fed with 44,000 spores had significantly higher proportion of foragers than bees from the other 3 treatments at worker ages of 7-18 days. In colony 17, High, middle and low proportions of foragers were found among bees fed with 44000, 4400, 440 and 0 spores at worker ages of 12-18 days. Points followed by the same letter at a certain worker age were not statistically different (G-test, P>0.05).
Discussion and Conclusion
This study clearly suggests genetic variation in behavioral response to Nosema infection (Fig.1). It is not clear why some bees are not responding to nosema infection behaviorally, because these bees also had large number of sprores in their midgut. Perhap these bees are resistant, not in the sense that they can reduce or eliminate Nosema spores, but rather their physiology is such that the i nfection causes no adverse effects on their behavior and physiology. The on-going program in selecting and breeding sensitive and resistant honey bee colonies will provide bee sources for our study of the mode of action of Nosema on bees. The bee breeding program will yield resistant stocks for beekeeping production in the future.
A concentration of 44,000 spores/bee is the optimal dosage (Fig. 2) for the future study of the effects of Nosema on honey bees.
A measurement of our stored bee haemolymph samples will give us a clue in the mode of action of Nosema on bees. If JH titers are high in the samples, it suggests that JH could be produced directly by Nosema, or possibly Nosema produce a substance that depresses JH degradation in the workers, resulting in JH accumulation. Alternatively, Nosema could produce a substance that enhances JH production by a host bee. If JH titers are low, Nosema might change the host physiology, resulting in workers being more sensitive to JH; Alternatively, Nosema could produce a novel factor, which causes the acceleration of behavioral and physiological development in workers. Future study will need to be followed to discern these hypotheses.
Acknowledgments
We thank Ray Ruiwu Wang, Candice Dusset, Joowook Park, and Micah Gill for their help in this study. This study is funded by USDA NRI grant (97-35302-5304).